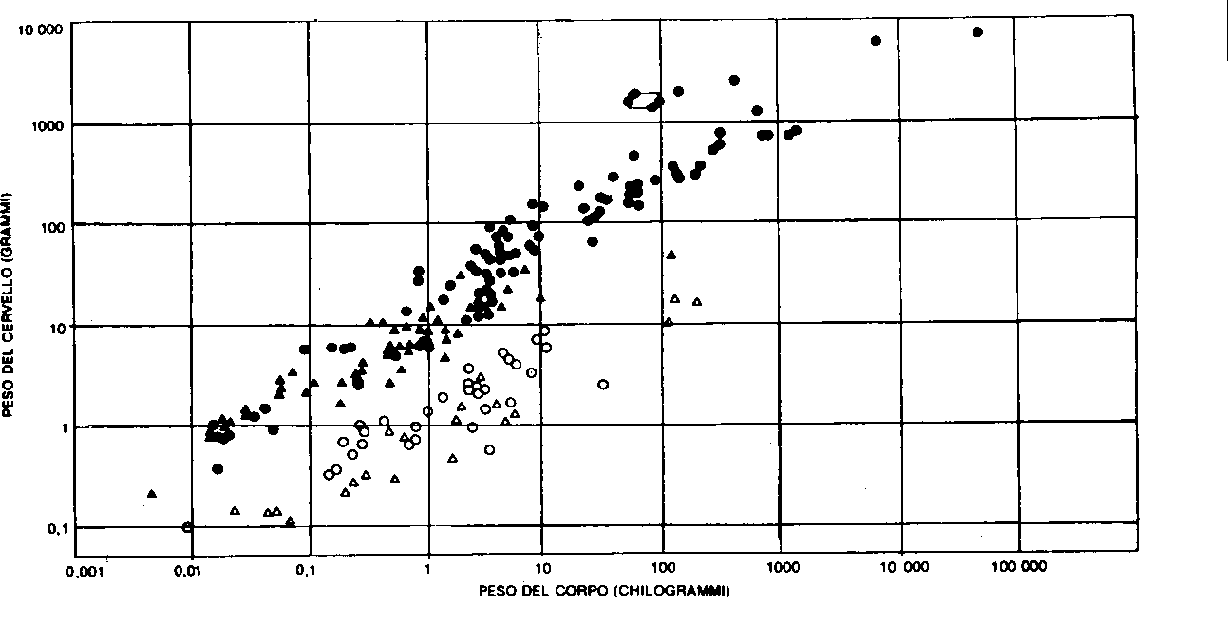
Paleoneurology and the origin of manby Mauro Mancia, Milan *In this paper I would like to present the contribution that neurophysiology has recently made to paleontological research. I will be making reference to the work of Jerison (1973, 1976), to the work of Jerison (1973, 1976), Holloway (1974), Leakey & Lewin (1977), Leroi-Gourhan (1965) and Eccles (1979). At the basis of this article is the concept that Figure I shows the ratio between body and the development of the brain and of its plasticity has permitted adaptations to environmental changes in such a way as to be a necessary step in any process of natural selection. Therefore the history of course of evolution that led to the human species is also and particularly the history of course of evolution of its brain and of the ever more complex operative circuits which have permitted Man to adapt to any change in his environment. An evaluation of this great development can be deduced from a study of the encephalization quotients (EQ) elaborated by Jerison (1973). This EQ indicates the ratio of the actual brain size to the expected one as given by the average for living mammals. It is expressed by the formula E = K.P 2/3, where E indicates brain weight, P body weight and K a constant which for mammals , can be considered equal to 0.12. Figure I shows the ratio between body and brain weight according to EQ.
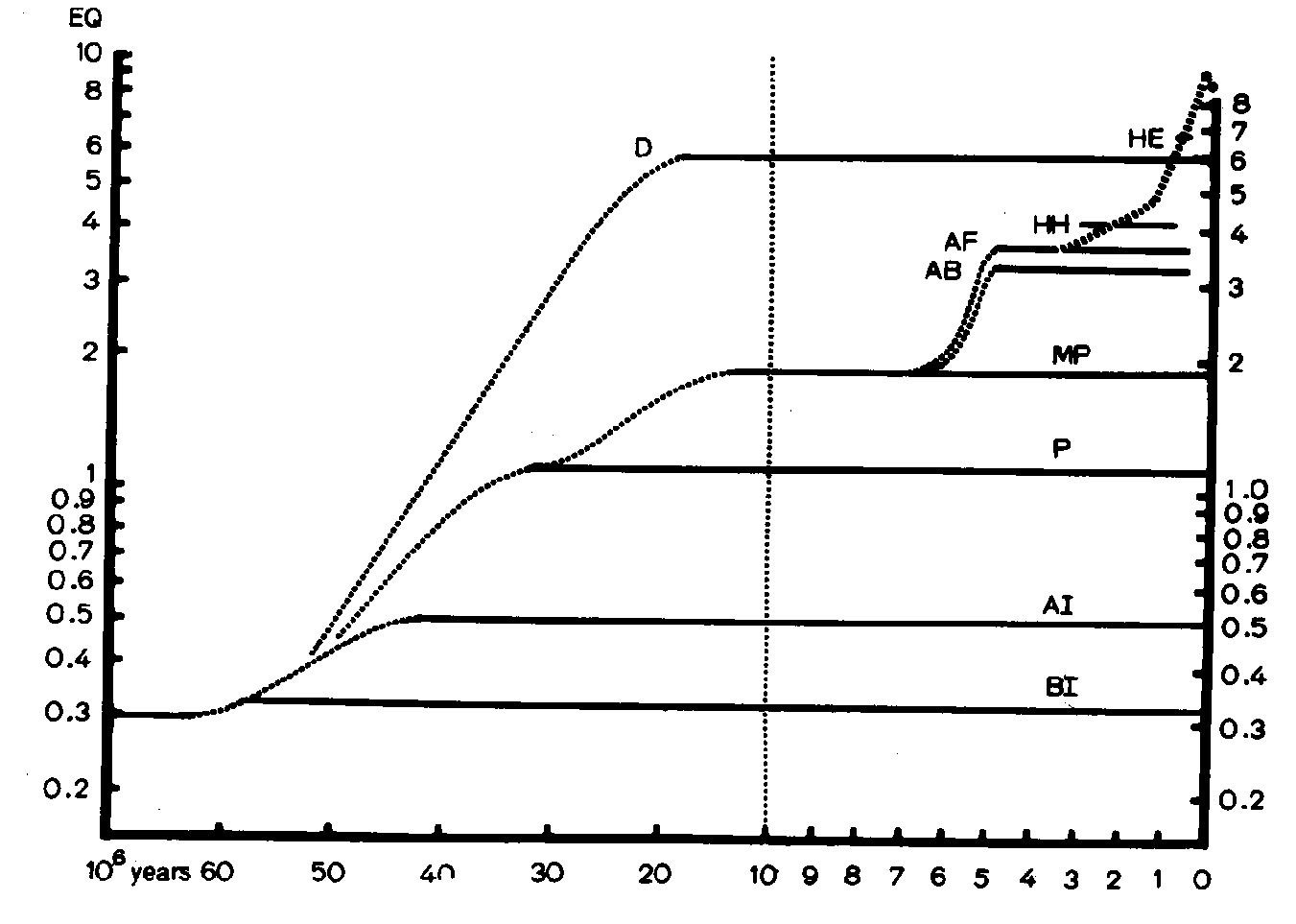
Two distinct goups are found: the higher represents mammals, primates and birds; the lower reptiles and skeleton fishes. The four linked points represent Man's EQ. All points lie along two curves whose slope is about 2/3 on the logarithmic coordinates, which means that the weight of the brain in different species varies with the body weight elevated to 2/3 according to the EQ formula. The study of the EQ has shown that mammals 60 million years ago had a EQ of 0.3 which has remained unvaried only for basal insectivores but has rapidly increased, starting 50 million years ago, for primates and dolphins. As can be seen in figure 2, dolphins began to increase their EQ, which reached the value of 6, as early as 30 million years ago, while prosimians remained at the value of 1.
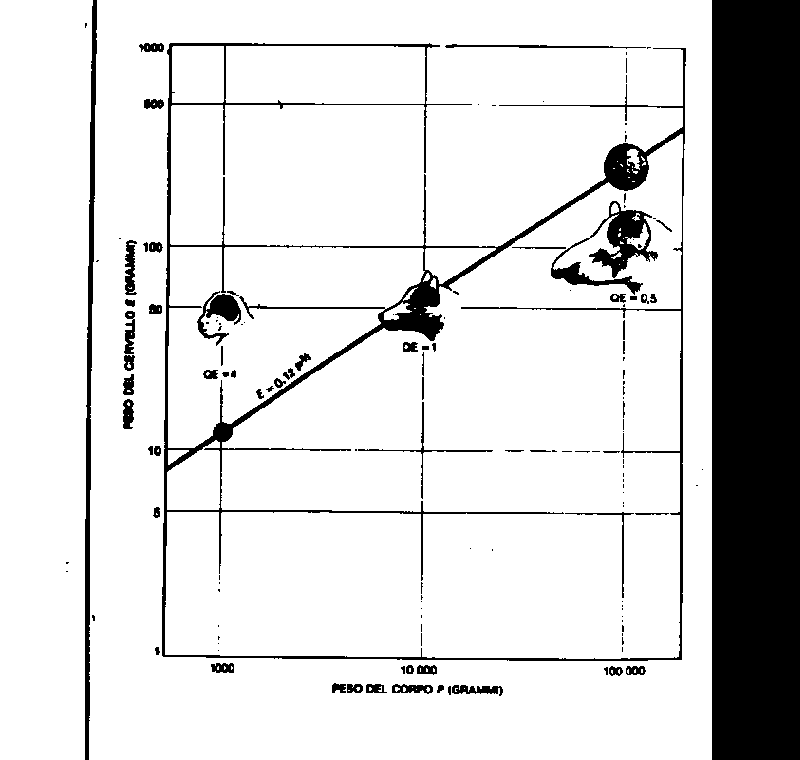
. The brain of Homo abilis developed from that of Australopithecus about 3 million years ago with a EQ of 4.2, while Homo erectus reached its EQ of 6.5 only 1 million years ago from which an EQ of 8.5 which characterizes Homo sapiens has developed. Figure 3 shows that from an evaluation of the EQ it is possible to distinguish three different points of the same curve for three different animals. The monkey, which has a brain four times larger (48 g.) than the presumed weight (12 gr.), has an EQ of 4. For a dog the ratio is only 1 and for a stupid animal like a tapir it is 0,5.
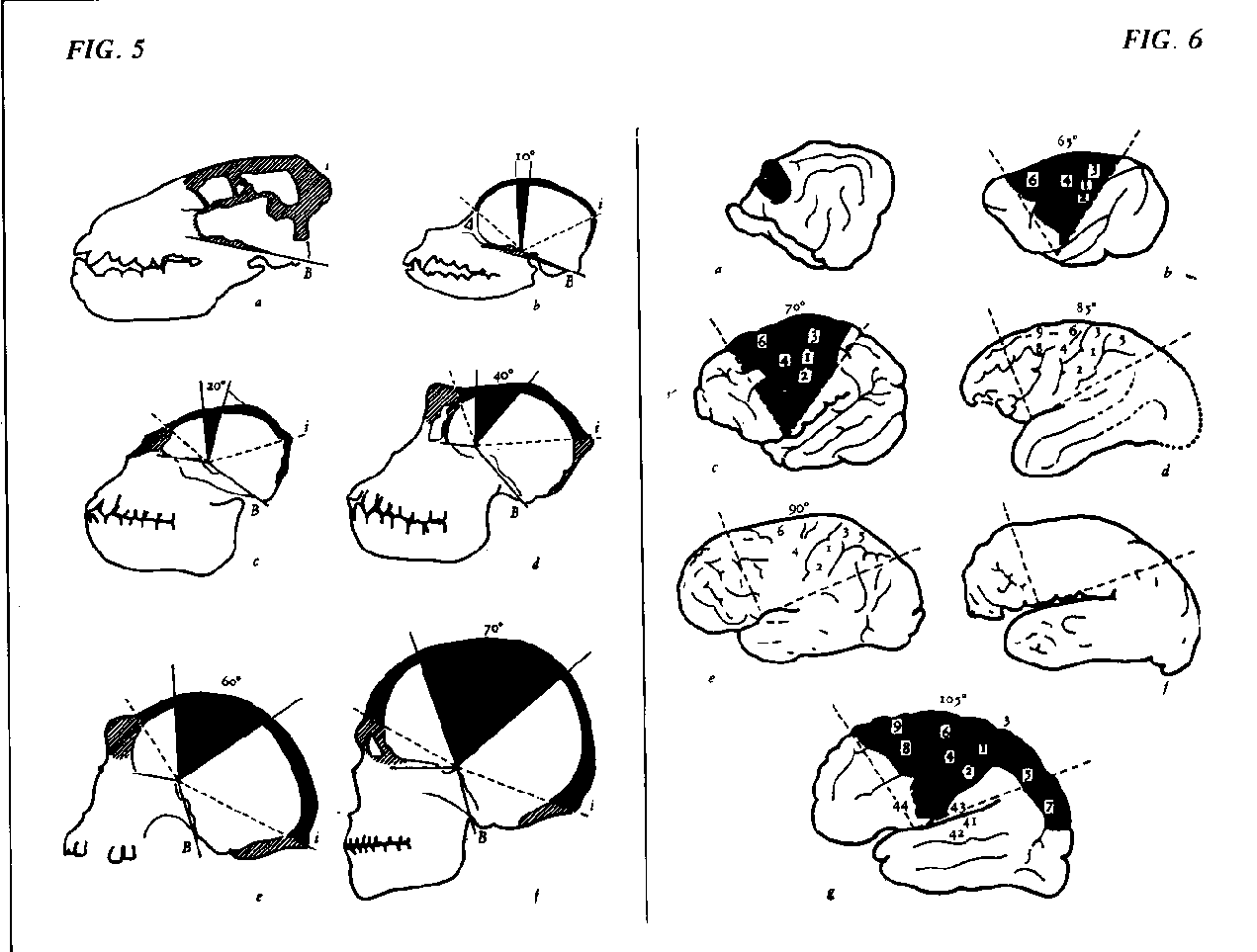
If we now look at the most significant contribution that neurophysiology has given to paleontology , we can see that it derives from particular techniques permitting the study of casts of fossil hominid brains, particularly of the Australopithecus discovered by Leakey in 1959. The evaluation of brain volume, shape and degree of corticalization thus achieved made it possible to advance an hypothesis on the origin of Man (Holloway, 1974).
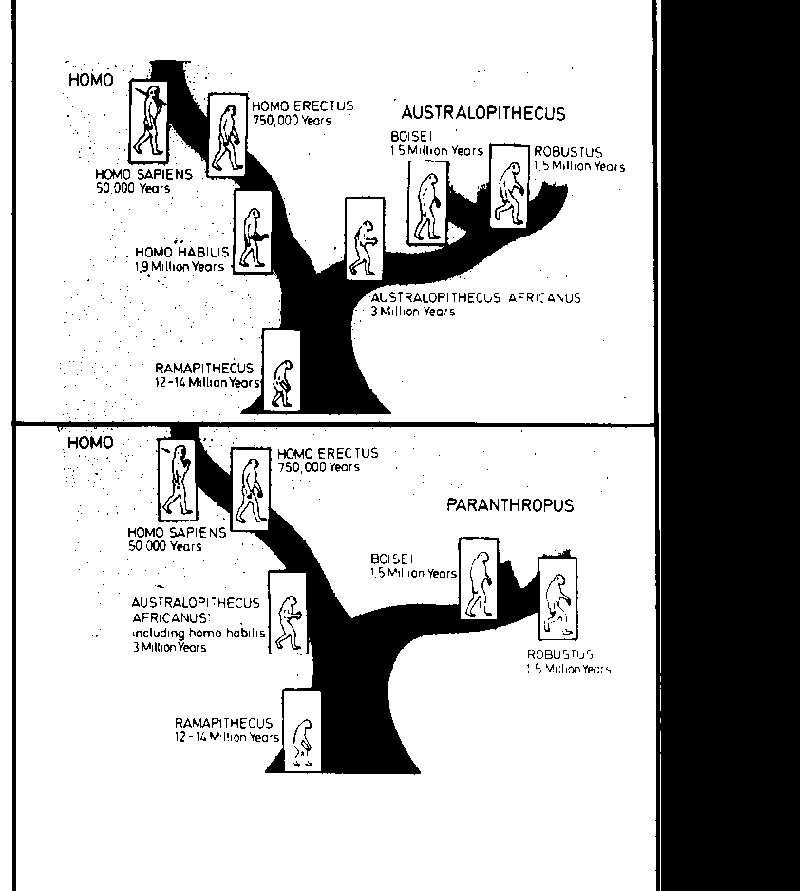
This hypothesis suggests that the common ancestor of man and monkey is the Driopitecus who lived in central and northern Europe, Asia and East Africa from 30 to 40 million years ago in the Cenozoic period. It is possible that Ramapithecus can already be considered to be a hominid on the basis of the study of the teeth and jaw, which are unfortunately the only fossils found. Ramapithecus had the same diffusion as Driopithecus and this seems to be a good reason for considering it to be the first link in a paleonthological chain eventually leading to Homo Sapiens.
Back in 1959, L.B.S. Leakey discovered in Tanganyika an Australopithecus which he called Zinianthropus Bosei, and which walked in an erect position, used tools and whose brain weighed 530 g. Successively, the discovery of another fossil in the coast of Lake Rodolfo, named E.R. 1470, whose skull dated at 2.8 million years ago, permitted the hypothesis that this hominid might represent Homo abilis whose brain weighed from 500 to 700 gr.
a) particular form of the tarsus, of the skeleton of the leg and of the pelvis;
Confirmation of this last feature is also found in Leakey's discovery (1976) of Australopithecus footprints fossilized in a layer of ash ejected 3.6 million years ago from the crater of a volcano
The considerable development of these completely new functions of the hand, the mouth and the organs of phonation now gives rise to development of the brain which becomes essentially human. Its volume increase progressively, and a widening of the medial fronto-temporoparietal region and a curvature of the cerebral vault are to be found. This latter develops side-by-side with a marked widening of the cranial vault which progressively opens out "like a fan" In Australopithecus this is incomparably greater than in the anthropomorphic ape and is destined to increase still further up to Home sapiens. The development of the cortical fan thus involves the areas of the associative brain, in other words that brain which neurophysiological research has related in the dominant hemisphere to language, gnosic and praxic functions, in other words to those superior functions which call for some degree of consciousness and permit some essential symbolic operations (Luria, 1962; Benton, 1965; see Bisiach ond others, 1978).
The contribution of paleoneurophysiology is thus to be identified in the discovery that new sensorimotor functions of adaptation to the environment and to locomotion correspond to significant modifications of cortical functions. Since such modifications have basically involved areas of the associative brain and their subcortical and inter-hemispheric connections, this means that symbolic and language functions, and hence all operations underlying creative processes have been conditioned over the millennia by evolutionary processes which primarily involved other functions and only in a secondary manner the specific development of determined cerebral areas.
Neurophysiological interpretation of paleonthological discoveries in fact bears evidence in favour that the change to erect locomotion goes hand in hand with an "avalanche" of events leading to a maximum of corticalization and a maximal development of the intellectual and creative capacities of Man.
Contingent to this paleontological consideration is the hypothesis (Popper and Eccles, 1977) that the liaison brain of the dominant hemisphere plays an essential part in the process of integrating information and activity "pattern" coming from the sensory world, as also in the operative mect~anisms underlying language functions. It is in these functions that we may find the foundations of a mind-brain theory which , though remaining dualistic in its formulation, aims nonetheless to identify a level of interaction in specialized areas of the dominant emisphere. I am referring here to the dualistic-interactionist theory put forward by Popper and Eccles (1977) and recently furhered by Eccles (1979). In this article we could hardly undertake a detailed analysis of this theorie which makes it possible to view the mind-brain relationship on an experimental footing. In another publication (Mancia, 1982) I have examined certain aspects on this theory. It will suffice here to recall that , according to the Eccles - Popper hypothesis the "liaison brain" is the seat of interaction between World 1 of physical objects and World 2 in which the self-conscious mind is able to "scan" and "read" the activity of each modular unit of the cortex (Szentagothai, 1978). By this operation the self-conscious mind can organize, select and integrate in a unitary operation the information which filters into the conscious mind through the interface represented by the liaison areas of the dominant hemisphere. This seems true also in ontogenetic development: the observation of a close correspondance and synchronization of the movements of the newborn with thelanguage of the adult (Condon & Sander,1974) suggests in fact the existence of a highly complex interactional system in which the motory behaviour of the newborn is guided andsynchronized by the adult who is speaking. Such a correlation indicates that the newborn child participates with a somethetic cerebral function (besides the acoustic and visual ones) in these basic neurobiological processees which underly the organization of its language. The synapses which the sensori-motor activity will form and activate in this process of maturation will involve those neurones of the liaison brain which make it possible which are later to govern the integration of sensori- motor functions proper to language. This process explains the child's interest in, and the meaning of, its innumerable repetitions of linguistic forms in forms proper to somestethic sensoriality, before it is able to transfer them into more-specific functions directly connected with speech. The language of the child will thus be affected by the neurophysiological organization of the liaison brain which it has achieved as a baby, an organization which can hardly fail to be influenced by the form and structure of the linguistic system, and by the culture to which it belongs and is developing. If these ontogenetic deductions are of any validity, they may then be taken to constitute a valid argument also in support of the hypothesis which attributes to the hand and to its sensori-motor to function the main responsability for the development of those cerebral areas which belong to the cortical fan, and which in the dominant hemisphere govern the symbolic and sensori-motor functions of language. The contribution of neurophysiological thought to paleontology is essentially this, and confirms that the development of the liaison brain is central in the mystery of human paleontology. They also throw light on the importance of particular sensori-motor conditions linked with locomotionin the differentiation , over millions of years, between specific brain functions which led form Ramapithecus to Australopithecus, and from these to the function of the human brain of today.
This paper reports some neurrophysiological contributions to paleontological research. The curves of the brain-body ratio and those of tile interactional encephalization quotients (EQ, given by E = K.P. 2/3 ) plotted against time up to the present age for various species, suggest that the EQ remains at 0.3 only for basal insectivores while dolphins has already reached the value of 630 million years ago and Home Erectus, who developed from Australopithecus reached an EQ of 7 only 1 million years before the present age.
Paleontological characteristics of the Australopithecus are presented which make it possible to consider this fossil a hominid with human capabilities including language and use of tools. NeurophysiologicaI evidence supports the paleontological hypotesis that bipedism and the release of hands from locomotory functions were the primary and basic element of human evolution.
The role played by the liaison brain in language and symbolization is discussed together with its implications for the dualist-interatioinist hypothesis put forward by Popper and Eccles.
In questo lavoro si riportano alcuni contributi neurofisiologici alla ricerca paleontologica.
Sono state presentate le caratteristiche paleontologiche dell'Australopiteco che permettono di considerare questo fossile un ominide con capacità umane, incluso il linguaggio e l'uso di utensili.
Il ruolo giocato dal cervello associativo in relazione al linguaggio e alla simbolizzazione viene in seguito discusso unitamente aile sue implicazioni con l'ipotesi dualista-interazionista elaborata da Eccles e Popper. REFERENCES
BENTON, A., Problemi di neuropsicologia, Ed. Univ., Firenze, 1965. * (Courtesy of “Primeval Sculpture”,1984, vol.1, number 1, and of Mauro Mancia, neurophysiologist, full.Professor of Human Physiology II, University of Milan, Italy, and full-member of SPI).
|